
seitonorika2016-1,2016年11月13日受理,ダウンロードPDF
(見本論文)カイコの翅分化における脱皮ホルモンの役割
大沢慶幸1,2,辻村秀信2
1.東京農工大学農学部蚕糸生物学科,〒183-8509東京都府中市幸町3-5-8
2.東京農工大学一般教育部,〒183-8509東京都府中市幸町3-5-8
責任著者:辻村秀信,tsujmr@seitonorika.jp,生徒の理科研究所,〒623-0342京都府綾部市金河内町
要旨:本研究では、カイコの翅の成虫分化におけるエクジソンの役割を明らかにした。まず、蛹期における翅の成虫分化の過程の組織学的変化を記載し、つづいて蛹化後様々な時期における中胸部結紮、結紮した個体への様々な濃度のβ-エクジソン注射により脱皮ホルモンの後翅の分化に対する効果を調べた。その結果、カイコ翅の発生における鱗毛の分化は蛹2日までに分泌されるエクジソンで十分であり、低濃度エクジソンにより促進され高濃度のエクジソンにより阻害される、成虫クチクラの分化は高濃度のエクジソンにより促進されることが分かった。
分野:生物 キーワード:カイコ、翅、エクジソン、変態
はじめに
鱗翅目昆虫の成長は、一連の幼虫脱皮、蛹脱皮、および成虫脱皮により区切られている(Wigglesworth 1972)。これらの脱皮は、前胸腺から分泌される脱皮ホルモン―エクジソン―の作用の下に、表皮組織が古いクチクラから分離し、新しいクチクラを分泌することによって起こるが、その際形成される表皮の性質は、幼若ホルモンの量によって調節される。つまり、幼若ホルモンが十分量存在する状態では幼虫の表皮が形成され、ほとんどない状態では蛹表皮が、全くない状態では成虫の表皮が形成される。
ところが、成虫の翅、足、触角などの表皮は、幼虫時には未分化な組織として成虫原基を形成し、一般の表皮とは異なる挙動を示す(Wigglesworth 1972)。すなわち、成虫原基の細胞は、幼虫期は緩やかに増殖を続けるだけで、幼虫脱皮に際しクチクラを分泌することはないが、蛹脱皮に近づくと著しい増殖を行い、反転して体表に位置するようになる。そして蛹クチクラを分泌し、蛹翅、肢、触角となる。その後蛹期において、蛹クチクラから離れ、その下に成虫のクチクラを分泌し、成虫の表皮に分化する。この変態期における成虫原基の劇的変化と、それに対するホルモン作用には強い興味がもたれ、多くの研究がなされてきた(総説としては、Ursprung and Nothiger, 1972)。しかしそのほとんどは、変態初期の成虫原基の増殖、反転や蛹クチクラの沈着に関するもので、成虫分化に関する研究は少ない。
翅原基は比較的均一で多量の細胞集団からなり、成虫分化において成虫クチクラ分泌の他に鱗毛を分化する。ホルモン調節の実験研究においては、中胸部の結紮により後翅への前胸腺の影響を除くことができる。
そこで本研究ではカイコの翅を用いて蛹期における成虫分化過程に対するホルモン調節を調べた。まず蛹期における翅の成虫分化の過程を組織学的に調べた。つぎに結紮が翅分化におよぼす影響と、蛹化直後に結紮した個体にβ-エクジソンを注射したときの後翅の分化をしらべ、翅の分化に対する脱皮ホルモンの効果を調べた。
材料と方法
カイコBombyx moriの実用品種、「紅白x同栄」および「群宝x秀玉」を飼育期間を通じ26℃、16L-8Dの条件で全令人工飼育(日本農産工K.K.、横浜)で飼育し、ほぼ同じ大きさの個体を選び、実験に用いた。この条件においては、蛹期間は約10日間であった。なお、以下の諸実験処理をした蛹は、同様の条件のもとで発生させた。
本実験では、蛹化後0~1時間を経過した蛹を蛹化後0日、蛹化後24時間を経過した蛹を蛹化後1日、蛹化後48時間を経過した蛹を蛹化後2日、以後同様に3日、4日・・・・9日と呼ぶことにした。また、羽化後1日の個体を成虫として取り扱った。各実験区においては、雌雄ほぼ同数とした。
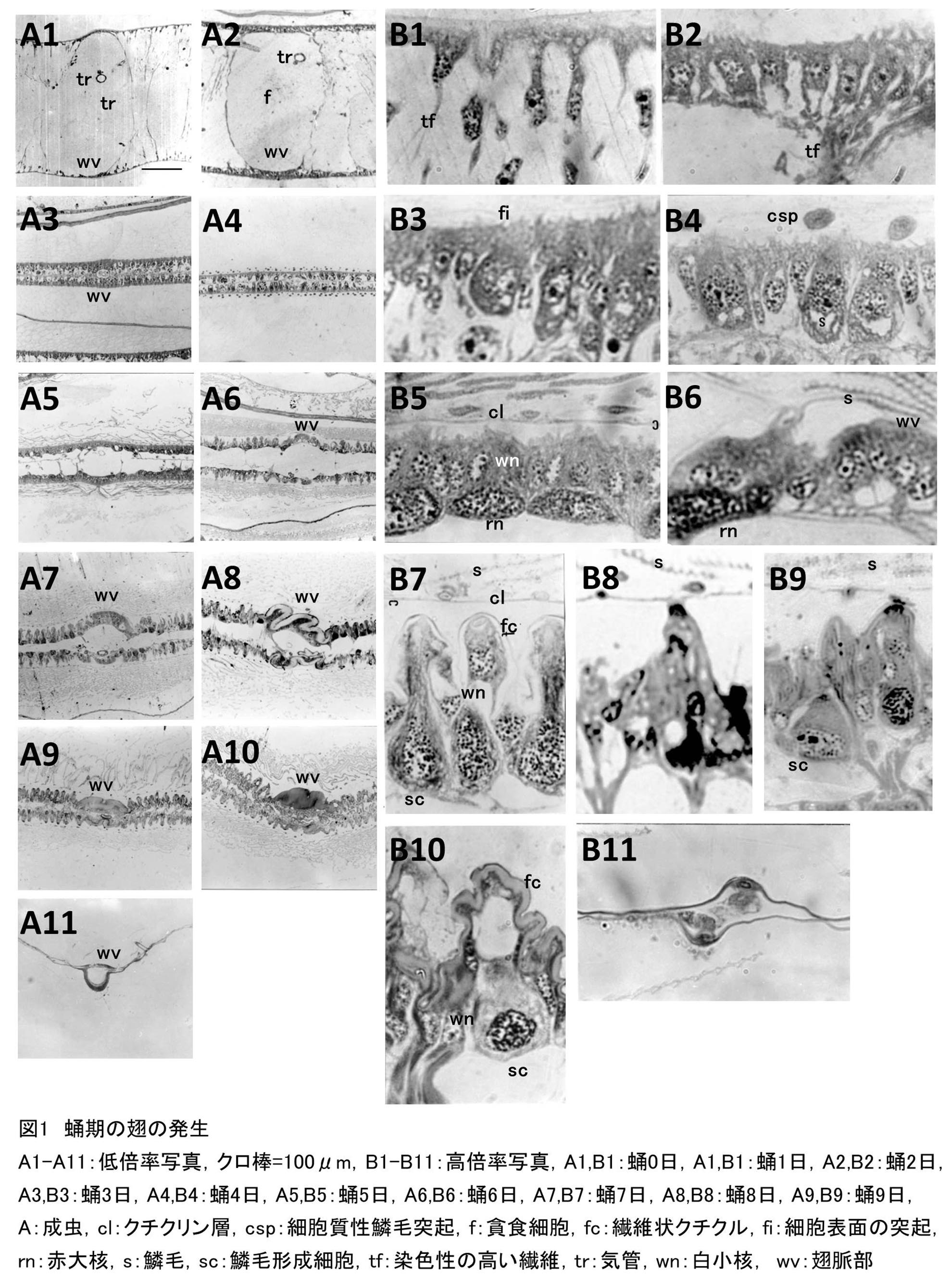
実験1 蛹期における翅の成虫発達の経過を知るために、蛹化後0日、1日、2日・・・・9日の蛹、および成虫の後翅を組織学的に調べた。実験は各区雌雄2頭ずつ行った。
実験2 翅の分化に及ぼす前胸腺ホルモンの影響を調べるために、蛹化後0,1,2,4,6,8日の蛹を2本の昆虫針をもちいて作製した松葉形ピンを用いて中胸部で結紮し、その頭側を切除し、切り口をパラフィンでふさいだ。この処理により、前胸腺は完全に除去されるが、後翅と腹部の体液的連絡は保たれる。結紮後、蛹期の日数が結紮前後あわせて10日間になるまで放置し、後翅の発達を組織学的に調べた。蛹化後、1,2,3,4日目については雌雄各4~6頭ずつ、0,6,8日については各2頭ずつ行った。
実験3 翅の分化に及ぼすβ-エクジソンの効果を調べるため、蛹化後0日の蛹を前述の方法で結紮し、その後2日間放置した後、減菌蒸留水にとかしたβ-エクジソン(ロート製薬Co.Ltd.、大阪)を、表面を70%アルコールで消毒した腹部第2環節に注射し、傷口をパラフィンで封じた。注射したβ-エクジソン量は、0.2、0.4、0.8、1.6、3.2、6.4および12.8μgである。注射した蛹は、その後10日間放置した後、後翅の発達を組織学的に調べた。実験は、各区について雌雄4-5頭ずつ行った。
各処理を受けた蛹や成虫から翅を切り出し、蒸留水(3)、90%アルコール(3)、フォルマリン(2)、酢酸(1)の割合で混合した液で1日間固定し、70%アルコール中に4℃で保存した。保存していた翅は、後翅先端部のみを切り取り、アルコールで脱水し、定法によりエポン包埋した。包埋試料からミクロトームを用いて1μmの切片を作製し、パラゴン染色液(Plysciences, Inc., Wormington)、または1%トルイジンブルー液で染色し、スライドグラスにエポン封入し光学顕微鏡で観察した。
結果
実験1 蛹期における翅の組織学的発生経過
正常に発育させた蛹の翅の成虫発達過程は次のようであった。また、雌雄の差異は認められなかった。
・蛹化後0日
蛹化後0日における翅の組織像は図1A1,B1に示した。翅の表裏細胞層は著しく離れ、内部は染色性の高い繊維が存在する部分と、全く存在しない部分に薄膜によって区切られている。貪食細胞が多数存在するのが観察されるが、形はまだ小さい。この時期にすでに翅脈部と他の部分は組織学的に区別できる。翅脈部以外の細胞層は薄く、また各細胞の形も不斉一である。染色性の違いや核の大きさには差異が認められず、細胞間の境界も不明瞭である。細胞の多くは、内部に向かって突起状に伸びており、そのいくつかは、内部の繊維とつながっている。翅脈部では、細胞層は他部より厚く、核は単層であるが、より集中している。翅脈部の内部には気管があり、染色性の高い繊維はない。細胞外表面は滑らかであり、染色性のない薄い透明な蛹クチクラに被われている。
・蛹化後1日
蛹化後1日における翅の組織像は図1A2,B2に示した。表裏細胞間の幅は、蛹化後0日より少し縮まり、個々の細胞および核は前日より大きくなる。細胞は立方体形となり、それが一列に整然と並び、表裏細胞層を形成している。細胞間の境界は明瞭となり、その内部には染色性のない核質を持つ小核が存在する(白小核)。翅脈部では細胞が気管を囲むようにゆるい弧を描いて配列しているが、細胞間の境界は明瞭ではなく、核のみが散在している。各細胞および核の染色性の違いはまだ見られない。表裏細胞層間の線維状物質、ファゴサイトの数、大きさなどは、蛹化後0日と目立った変化はない。細胞層の外表面は蛹クチクラと分離し、蛹クチクラは前日より厚くなり、染色性は高くなっている。
・蛹化後2日
蛹化後2日における翅の組織像は図1A3,B3に示した。表層細胞層間の幅は著しく狭くなる。細胞は著しく成長し、翅原基内部に向けて伸長する。ある間隔をおいて、ふつうの真皮細胞とは形、大きさの異なる大きな楕円形をした鱗毛形成細胞と思われる細胞があらわれる。この細胞は、大きな核を持ち、細胞表面から短い細胞質性の突起を出しているものもある。鱗毛形成細胞以外の細胞は小さく、核の配列も不規則である。その細胞の表面からは、小さく細い突起が多数出ているが、これは蛹化後4日まで観察される。表裏細胞間は層状の物質によって連絡され、その物質は翅脈部では表裏細胞層の基底部を気管を囲むように結んでいた。貪食細胞は細胞間、翅脈部などに数多く存在し、前日までのものより少し大きくなっている。
・蛹化後3日
組織像は図1A4,B4に示した。鱗毛形成細胞は前日より成長し、その形はフラスコ形となる。細胞質、核とも他の真皮細胞とは染色性が強く、明瞭に区別できる。つまり、鱗毛形成細胞はパラゴン染色液により赤く染まり、その核は赤く濃染するが(赤大核)、他の細胞は紫に染まり、その核は染色されない(白小核)。鱗毛形成細胞の先端からは細胞質性の突起が伸び(細胞質性鱗毛突起)、翅の表面にはその楕円状、円状の断面が列をなして並んでいるのが観察される。したがって、この突起は屈曲しているものと想像される。鱗毛形成細胞の内部には、染色されない空胞が存在する。翅脈部、貪食細胞などには変化は見られない。
・蛹化後4日
蛹化後4日における翅の組織像は図1A5,B5に示した。組織の様相は一変し、内部に向かって伸びていた細胞はすべて横並びに密着して層状となり、細胞層の内表面は平滑となる。表裏細胞層の間隔がひろがるとともに、組織全体の厚みも増す。鱗毛形成細胞はさらにフラスコ形に近づき、その底部に赤大核をもつ。他の真皮細胞は白小核を持ち、細胞層は表面側に白小核を持つ普通の真皮細胞、基底部側に赤大核が並んだ様相を呈する。またこの時期に初めて鱗毛が観察される。しかし鱗毛の量は少なくその断面も凹凸がなく平板である。また内部にはまだ細胞質が残存しているらしく、青色に染色される。細胞外表面に、細胞層から狭い間隔をおいて直線的に伸びる薄層が観察される。この層はエピクチクルの一部であるクチクリン層と思われる。この層と細胞層との間の透明で染色されない部分は、タンパク質性クチクルと思われる。クチクリン層は、鱗毛形成細胞の突起の先端を結ぶようにして伸びている。3日目まで表裏細胞層を連絡していた線維性物質は減少し、また貪食細胞は表裏細胞層の間隔中に散在するようになる。
・蛹化後5日
この時期の組織像は図1A6,B6に示した。鱗毛形成細胞をはじめとして、各細胞は外に向かって凸状となり、細胞層外表面には著しい凹凸ができる。鱗毛形成細胞は、はっきりとしたフラスコ形を呈するようになり、その底部に赤大核、またその頸部にソケット細胞のものと思われる白小核を持つ。細胞層内表面はゆるやかな弧を描くように整っているのが観察される。クチクリン層は蛹化後4日より一層明瞭となり、鱗毛形成細胞の突起の先端部を結ぶようにして直線的に伸びる。また、クチクリン層に一定の間隔をおいて黒点が存在したが、これは鱗毛が出ている箇所と推察される。クチクリン層はこのあと変化なく、蛹化後9日目まで同様に観察された。この時期において、細胞外表面の凹凸部に、パラゴン染色液で薄桃色に染色される新しいクチクラ層が沈着し始めているのが観察された。この層は繊維状クチクルもしくはエンドクリクルと呼ばれる層である。鱗毛は蛹化後4日より量的に少し増え、その表面には縦助と呼ばれる凹凸の形成が認められた。また、鱗毛の染色性は落ち、細胞質が消失しつつあるものと思われる。表裏細胞層を連絡していた線維状物質はほとんど消失し、貪食細胞も少数となる。
・蛹化後6日
蛹化後6日における翅の組織像は図1A7,B7に示した。表裏細胞層間の間隔は翅脈部を除いて狭くなる。各細胞は縦長に発達し、各細胞、核とも境界が明瞭となり、底部に赤大核をもつフラスコ形の鱗毛形成細胞、その頸部を取り巻く白小核を持ったソケット細胞がはっきりと認識できるようになる。翅脈部は細胞の境界が明瞭でなく、白小核、赤大核が混在しているのが観察される。繊維状クチクルの沈着は進み、細胞層外表面の凹凸を覆うようになる。鱗毛の存在する層も厚くなり、鱗毛は表面のクチクラのみが染色され、内部の細胞質はほぼ消失している。表裏細胞層間を連絡していた繊維性物質は消失するが、貪食細胞はまだ少数見られる。
・蛹化後7日
蛹化後7日における翅の組織像は図1A8,B8に示した。細胞の形は不斉一となり、表裏細胞層の内壁も平滑ではなくなる。翅脈部は細胞層の外表面の凹凸が激しくなり、また、細胞層も薄くなり、その中に核が散在するようになる。前日に比べ、繊維状クチクルの沈着は厚くなり、特に翅脈部には著しい量の繊維状クチクルが沈着されているのが観察される。鱗毛量は前日と変わらず、この後も増加することはなかった。しかし、形態的には正常なもののほかに星形、楕円形などの形の鱗毛がところどころに散在していた。表裏細胞層間には、一部の細胞から伸びてできたと思われる、染色性の高い繊維性物質が、ところどころ両細胞層を連絡しているのが観察された。貪食細胞はこの時期には見られなかった。
・蛹化後8日
蛹化後8日における組織像は図1A9,B9に示した。表裏細胞層間の間隔はさらに狭くなり、前日観察された線維性物質は随所に現れ、発達し、またその中には染色性の高い物質が散在しているのが観察された。
・蛹化後9日
組織像は図1A10,B10に示した。表層細胞を連絡する線維性物質がより発達する。また中にある染色性の高い物質も大きくなり数も増える。細胞層のところどころに間隙が観察され、成虫化に向け個々の細胞の細胞質が縮小または退化しているものと思われる。
・成虫
成虫における翅の組織像は図1A11,B11に示した。表裏2層のクチクラが直線的に伸び、その間隔は著しく狭くなっている。各細胞の細胞質はほぼ消失し、個々の細胞は全く識別できなくなる。鱗毛を突出している部位はクチクラが山型にふくらみ、その基部に鱗毛形成細胞のものと思われる核が存在する。翅脈部は半円形に大きく膨らみ、クチクラも沈着している。ここでは厚い細胞層がクチクラの内壁に沿って認められ、その内部には核が散在しているのが観察される。翅脈内部には気管や神経が存在する。単位面積当たりの鱗毛量は少ないが、これは翅が伸展したためであると思われる。
実験2 中胸部結紮の翅分化に対する影響
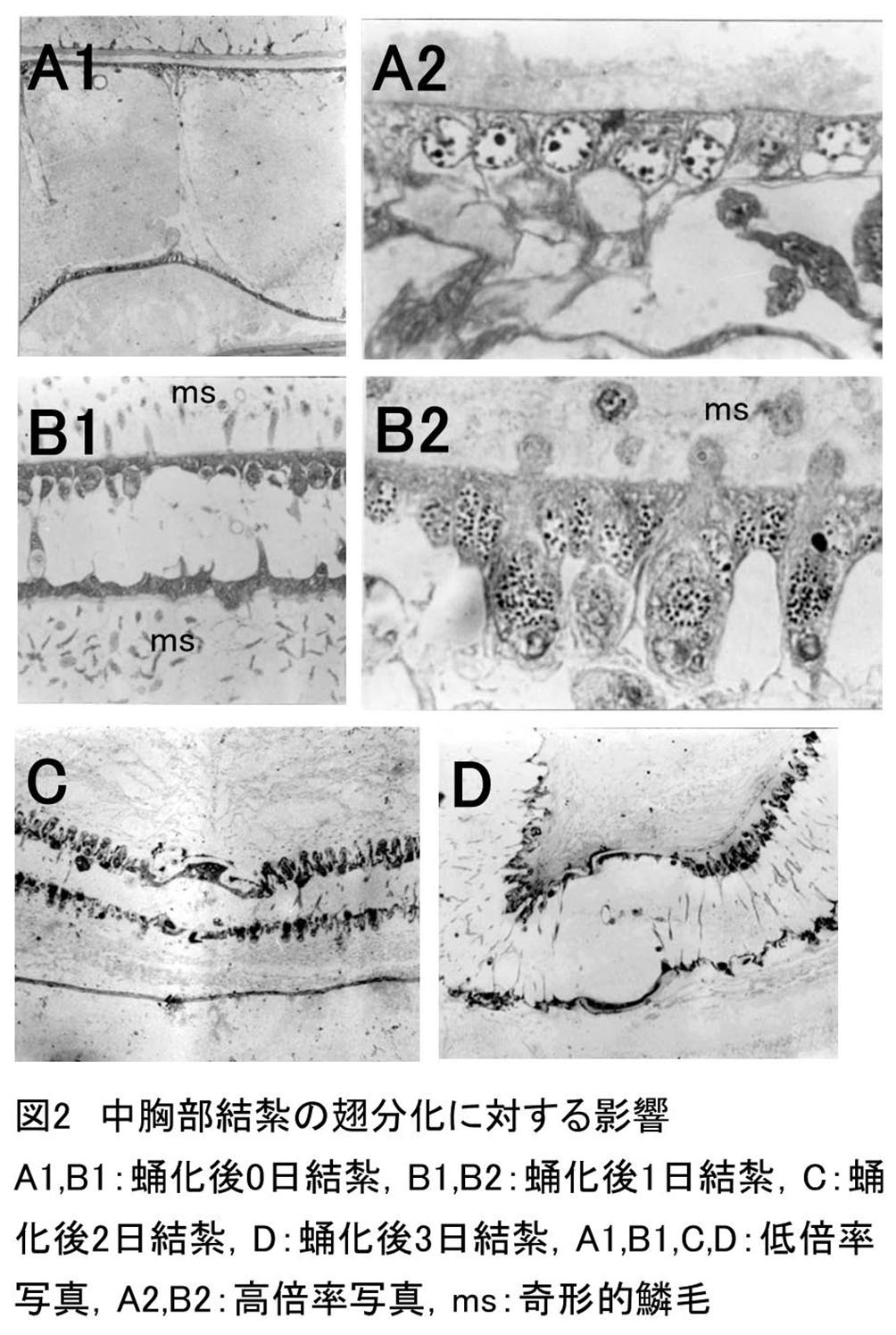
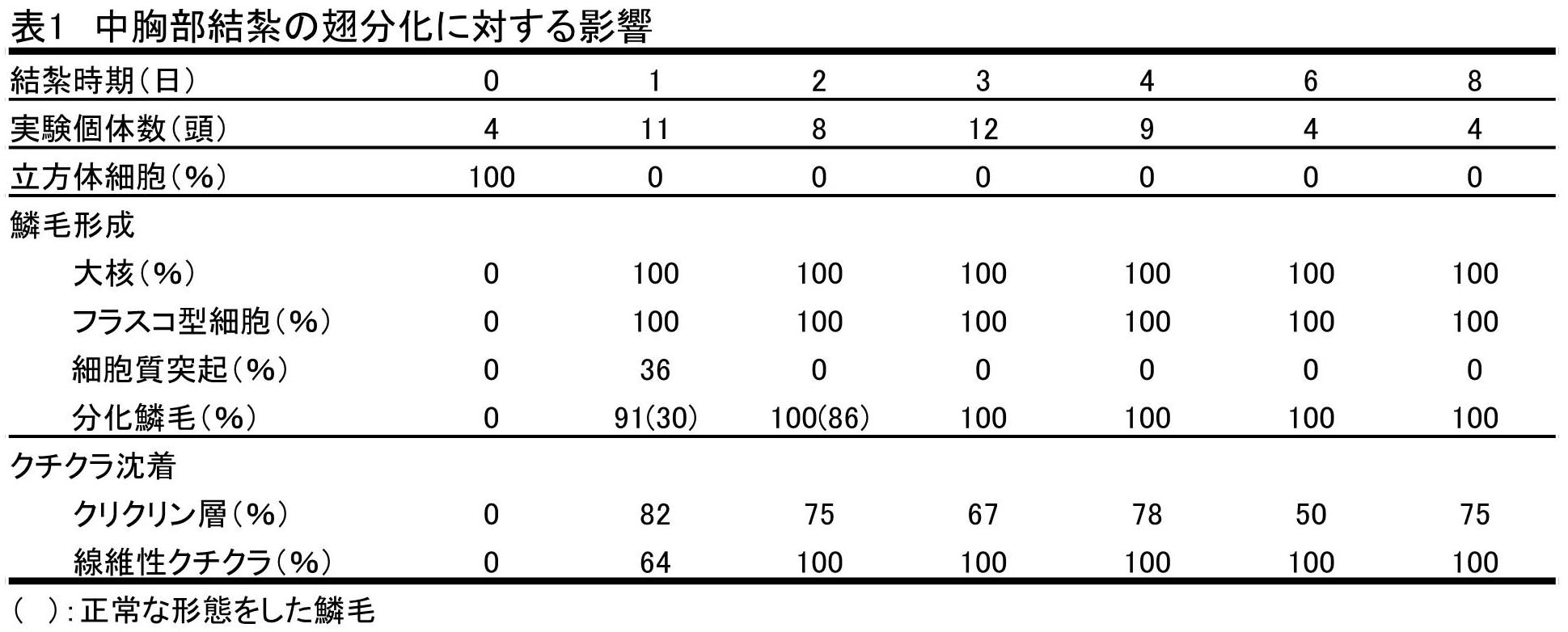
蛹化後、様々な日にちをおいて中胸部を結紮した後、発生させた場合の後翅の組織像をまとめると表1 のようであった。
 ・蛹化後0日結紮
・蛹化後0日結紮
すべての個体において成虫分化の兆候は見られなかった(図2A1B1)。すなわち、個々の細胞は立方体形になり、それらが一列に並んで表裏細胞層を形成していた。また、その核は均一であり、赤大核の分化は認められなかった。
一方、蛹外皮は蛹化0日に較べ肥厚化して着色し、蛹クチクラ特有のキノンタンニングが認められた。この蛹外皮は細胞層と分離していたが、その程度は小さかった。また細胞外表面は平滑であり、成虫クチクラ分泌の兆候は全く見られなかった。
・蛹化後1日結紮
図2B1B2はこの区において観察された組織像の一部である。鱗毛形成細胞の分化を示す赤大核およびフラスコ形細胞は、全個体において観察された。鱗毛の分化に関しては、分化した鱗毛は91%個体において見られたが、正常な形態をした鱗毛は36%個体にしか観察されなかった。他の70%個体においては、鱗毛が扁平にならず、円形、楕円形のままの奇形的鱗毛であった。このうち30%個体においては、鱗毛内部に細胞質が残存し、また同時に細胞質性鱗毛突起も観察された(図2B1)。残りの40%個体においては、鱗毛内の細胞質はすでに退化しつつあり、奇形ではあるが鱗毛として完成された形態となっていた(図2B2)。
鱗毛を分化した個体について形成した鱗毛量をみると、正常に発生させた個体とほぼ同量(標準量)の鱗毛を形成したものは少数(30%個体)であり、他の70%個体は少量の鱗毛しか形成しなかった。このうち40%個体は奇形的鱗毛として完全に分化したものだった。
クチクラ沈着に関しては、クチクリン層は82%個体において観察され、また繊維状クチクルは64%個体において観察された。繊維状クチクルを沈着していない個体のうち27%個体は鱗毛の分化途中の組織像を示した。36%個体について、翅内部に貪食細胞が観察された。
・蛹化後2日~8日結紮
これらの実験区における典型的な組織像は図2C,Dに示した。鱗毛形成細胞はすべての個体において観察され、また鱗毛の分化もすべての個体に見られた。蛹化後1日結紮区において見られた奇形の鱗毛は、蛹化後2日結紮区において1個体見られただけで、他はすべて正常な形態をした鱗毛が分化した。鱗毛量に関しては、ほとんどの個体において標準量の鱗毛が形成された。
クチクラ沈着に関しては、クチクリン層の形成は各区において50~80%観察され、繊維状クチクルの沈着はすべての個体において観察された。
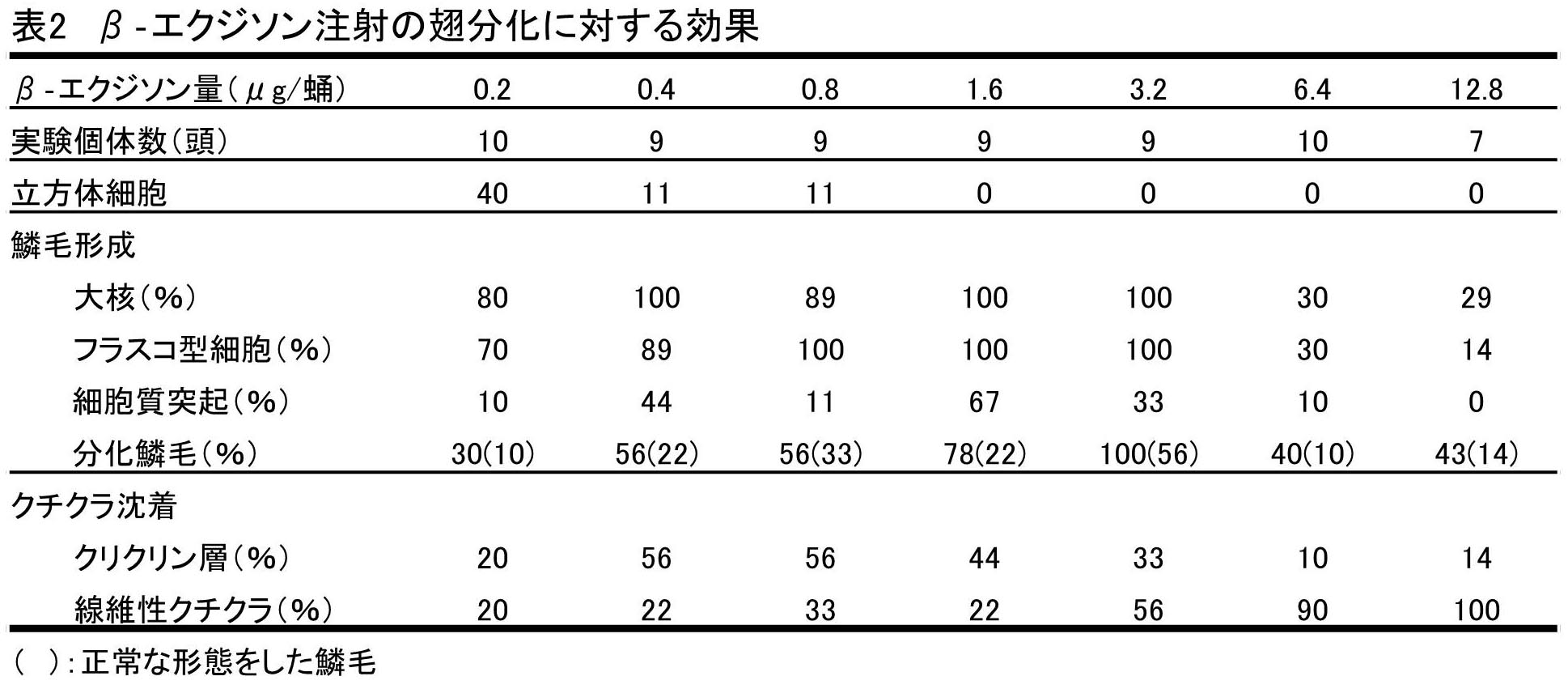
実験3 β-エクジソンの翅分化に対する効果
蛹化直後結紮し、2日間放置した後、各種濃度のβ-エクジソンを注射し発生させた時の組織像をまとめると、表2のようであった。結紮後2日間放置してからエクジソンを注射した理由は、予備実験で、蛹化直後に結紮し、結紮後の様々な時期の組織像を観察した結果、結紮後2日目にすでに実験2における蛹化後0日結紮10日後観察区と同様の組織像が観察され、翅分化に対するエクジソンの効果は結紮2日後の蛹にエクジソン注射することにより適切に見ることができると判断したからである。実験の結果はつぎのようであった。
・0.2μg注射
この区においては、鱗毛形成へ向かう様々な段階の組織像を持つ個体が得られた。すなわち赤大核を基底部に持つフラスコ形細胞は70%個体に観察された。一方、フラスコ形細胞は見られないが、立方体細胞と、その中に赤大核の分化のみが認められるものが10%個体あった。残りの20%個体においては、フラスコ形細胞、赤大核の分化は認められず、細胞は立方体型で均一であったが、この場合にも蛹化後0日結紮より細胞は大きくなり、エクジソンの効果が認められた。
鱗毛の分化に関しては、30%個体において分化した鱗毛が観察された。このうち10%個体においては、正常な形態をした完全に分化した鱗毛が存在したが、他の20%個体においては、多くの鱗毛はまだ十分扁平になっておらず、内部にはまだ細胞質が残っていた。鱗毛形成の初期段階である細胞質性鱗毛突起は10%個体について観察されたが、この個体はまだ鱗毛を分化していなかった。
クチクラ沈着に関しては、大部分の個体において認められなかった。ただ鱗毛を形成した個体においてのみクチクリン層および繊維状クチクルの沈着が認められた。
・0.4~3.2μg注射区
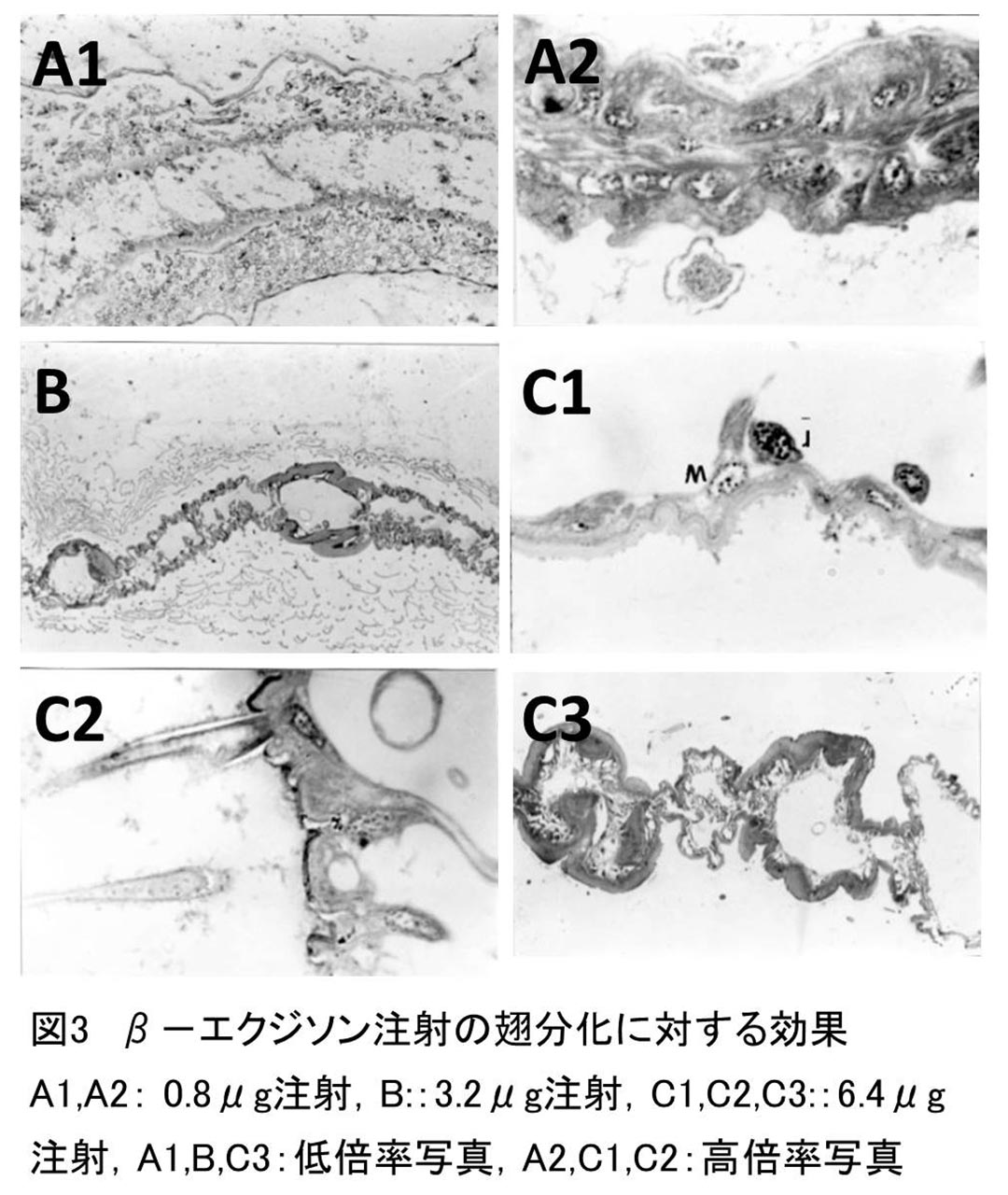
図3A1,A2,Bは、これらの区において観察される組織像の一部である。0.4, 0.8μg注射区においてフラスコ形細胞を分化していないものが各11%個体ずつ観察されたが、他のすべての個体においては、基底部に赤大核をもつフラスコ形の鱗毛形成細胞が分化した。また、3.2μg注射区の22%個体においては、染色性の低い小型の核(白小核)、鱗毛形成細胞特有の大きく染色性の高い核(赤大核)の他に、大きさは白小核とほぼ同じだが、染色性は高い核(赤小核)が散在しているのが認められた。
鱗毛形成に対する0.4~3.2μgエクジソン濃度の効果は、鱗毛の分化という観点から見てみると、鱗毛の分化は濃度が上がるにつれより多くの個体に観察され、3.2μg注射区においては、全個体が鱗毛を分化していた。しかしどの区においても形成された鱗毛の分化の程度や形態は様々であり、完全に分化し、正常な形態をした鱗毛をもつものは、どの濃度区においても鱗毛を形成した個体数の約1/2しかなかった(図3B)。他の個体においては、扁平にならず、断面が円形や楕円形のままで縦助が形成され、細胞質も退化、消失してしまった奇形的鱗毛や、同様の形態で鱗毛表面や細胞層表面にクチクリン層が形成されているが、鱗毛内部にはまだ細胞質が存在している個体、および細胞質が退化過程にある個体が観察された(図3A1,A2)。
しかし、鱗毛形成量という観点からとらえると、標準量形成したものは0.4μg注射区で33%個体、0.8μg注射区で44%個体といったん上昇するが、0.8μg注射区以降は、1.6μg注射区で22%個体、3.2μg注射区で33%個体と著しく低くなった。すなわち3.2μg注射区では鱗毛を形成した個体の多くは、少量の鱗毛しか形成しなかった。また、鱗毛の形成過程にみられる細胞質性鱗毛突起が多くの個体でみられた。すなわち、0.4μg注射区に44%個体、1.6μg注射区に67%個体、3.2μg注射区に38%個体、0.8μg注射区に11%個体で見られ、そのいくつかでは細胞質の詰まった鱗毛、または奇形的鱗毛と同時に観察された。
クチクラ沈着に関しては、クチクリン層は0.4μg、0.8μg注射区にそれぞれ56%個体と最も多く観察されたが、それ以降、濃度が上がるにつれ徐々に減少していった。一方、繊維状クチクルの沈着が認められた個体は、0.4~1.6μg注射区においては22%または33%個体と少数であったが、3.2μg注射区においては56%個体と増加した。
・6.4, 12.8μg注射区
これらの注釈ではおもに図2C1,C2,C3のような組織像が得られた。これらの区においては、鱗毛形成細胞の分化に大きな変化が見られた。すなわち赤大核やフラスコ形細胞は、6.4、12.8μg注射区では少数の個体でしか観察されなかった。しかし、6.4μg注射区に70%個体、12.8μg注射区に70%個体、白小核と混ざって前述した赤小核が多数観察された(図2C1)。また6.4μg注射区の1個体においては、この赤小核をもつ細胞から鱗毛が突出している像が観察された(図2C2)。しかしこの細胞は、鱗毛形成細胞に典型的なフラスコ形はしていなかった。
鱗毛の分化に関しては、分化した鱗毛が形成された個体は、6.4μg注射区で40%個体、12.8μg注射区で30%個体と少数であり、しかも形成された場合にも、ほとんどの個体では奇形的鱗毛であり、形成量も少量であった。鱗毛が少量形成された個体の、またはほとんど形成されなかった個体の大部分は前期の赤大核やフラスコ形細胞はほとんどなく、赤小核を持つ個体であった(図2C2)。
一方、クチクラ沈着に関しては、クチクリン層は、6.4、12.8μg注射区とも1個体にしか見られなかった。しかし繊維状クチクルは6.4μg注射区の1個体を除き、すべての個体において著しい沈着が見られた。
考察
1)翅の成虫分化の組織学的経過
本研究で明らかになった蛹期における翅の成虫分化を要約すると、次のようになる。
蛹化後0日においては、翅組織は様々な形をした小細胞の層よりなり、その外表面には薄く透明な蛹クチクラに被われている。蛹化後1日になると、個々の細胞はより大きくなって、立方体型となり、横に一列に並び、真皮細胞層を形成する。また蛹外皮はキノンタンニングにより着色し、表裏細胞層よりアポリシスする。蛹化後2日には、ふつうの真皮細胞の他に大きな核を持つ鱗毛形成細胞が多数分化する。蛹化後3日目には鱗毛形成細胞は1本の太い細胞質性の突起を外に突き出す。蛹化後4日になると、突起は扁平になり、表面にクチクラを沈着し始める。またこの時期には、細胞層の表面にクチクリン層が沈着する。ついで鱗毛内部の細胞質は退化し始め、鱗毛は表面にクチクラを残すのみとなり、鱗毛分化は完了する。一方、この時、表裏細胞層外表面には、繊維状クチクルが沈着し始める(蛹化後5日)。その後、翅脈部をはじめ、表裏細胞層の外側に繊維状クチクルが厚く沈着してゆく。羽化前日には、各細胞は羽化に向けて退化、収縮してゆく徴候を示す。そして羽化とともに翅は著しく進展し、成虫翅として伸展される。
以上の結果は、同じカイコを用いた池田(1913)の報告より各事項とも少々過程が早かったが、ほぼ同様であった。
2)蛹期に分泌される前胸腺ホルモンの翅分化に対する役割
鱗毛形成細胞は、蛹化後0日結紮区においては全く見られなかったが、蛹化後1日以降の結紮区ではすべての個体において見られた。ところが分化した鱗毛については、蛹化後1日結紮区で大部分の個体について見られたがその多くは、量的に少量しか形成されず、また奇形的鱗毛が多かった。そして蛹化後2日結紮区において初めて、大部分の個体に正常な形態をした標準量の鱗毛が形成された。以上のことにより、鱗毛形成細胞の分化には蛹化後0日から蛹化後1日までに分泌される前胸腺ホルモンで十分であることがわかる。一方、完成された鱗毛の分化には、蛹化後2日までのホルモンが必要であるといえよう。
蛹化後0日結紮区においては、蛹クチクラのタンニングと細胞層からの分離が見られたが、成虫クチクラの沈着は認められなかった。蛹化後1日結紮区においては、64%個体、蛹化後2日以降結紮区においてはすべての個体において成虫の繊維状クチクルの沈着が認められた。以上の結果より繊維状クチクル(エンドクチクラ)の沈着は、蛹化後2日までに分泌された前胸腺ホルモンで十分であるといえよう。
以上から蛹化後2日までに前胸腺から分泌されるエクジソンは十分にその後の順序だった成虫分化を進めることができるといえよう。この事実を蛹期における体内エクジソン量の経日的変化と考え合わせると非常に興味深い。カイコの蛹化直後の体内エクジソン量はほとんど0であり、その後急激に増加し、蛹化後2日目にβ-エクジソンに換算して1頭当たり役0.8μgにまで達する。そしてその後減少し、5日目で再び0となる山形のカーブを描く(Hanaoka and Ohnishi, 1974)。この報告と今回の実験結果を考え合わせると、翅の成虫分化には蛹化後濃度が上昇し2日目にピークに達するまでのエクジソンが重要であり、その後5日目までの減少過程に存在するエクジソンは必要不可欠なものではないといえる。
3)翅分化に対するβ-エクジソンの適正濃度
赤大核をもちフラスコ形をした典型的な鱗毛形成細胞は、0.2μg注射区においては70%個体、0.4μg注射区から3.2μg注射区においては大部分の個体において観察された。ところが、6.4μgおよび12.8μg注射区においては各30%個体および10%個体にしか観察されなかった。
また鱗毛は、0.2μg注射区においては30%個体で分化し、それ以降濃度が上がるにつれ増加し、3.2μg注射区では全個体に鱗毛の分化が観察された。しかし、6.4μg, 12.8μg注射区においては各60%個体および43%個体と鱗毛を分化した個体は、鱗毛形成細胞を分化した個体同様少なかった。
クチクリン層は0.2μ注射区においては50%個体に観察された。しかし1.6μg注射区以降は徐々に減少し、6.4μg, 12.8μg注射区においては各1個体ずつにしか観察されなかった。
一方、繊維状クチクルは、0.2μgから1.6μgまでの注射区においては20%~30%個体にしか観察されなかったが、3.2μg注射区以降上昇し、12.8μg注射区においてはすべての個体において沈着が認められた。
以上の結果により、各区の全個体数の50%以上に認められた場合を臨界点とすると、鱗毛形成細胞の分化は0.2~3.2μg、鱗毛の分化は0.4~3.2μg、クチクリン層は0.4~0.8μg、繊維状クチクルの沈着は3.2μg以上のエクジソンが有効であるといえる。これらの事実は、一連の翅の成虫分化の構成要素である鱗毛形成細胞の分化、鱗毛の分化、および繊維状クチクルの沈着は、それぞれ異なるエクジソン濃度により発現されることを示している。また、鱗毛形成においても、鱗毛形成細胞の分化とそれ以後の鱗毛分化の諸段階は、一つの連続した過程ではなく、各々異なるエクジソン濃度を必要とすること示している。
これと同様なことが他の成虫原基の研究でも明らかにされている。すなわち、同じ鱗翅目昆虫P.interpuntellaの翅の成虫原基は、in vitroにおいて100%の反転を引き起こすのに、0.2μg/ml β-エクジソンで15時間処理しなければならないが、100%のクチクル沈着を誘導するには、同濃度で24時間処理することが必要である(Butkowski and Oberlander 1973, 1974)。また、Agui and Fukaya(1973)もM.brassicaeの翅原基において同様の事実を報告している。
したがって、鱗翅目昆虫の変態において、成虫原基に起こる諸過程は、ある程度独立した過程であり、それぞれ異なる濃度のエクジソンにより調節されていると考えられる。蛹におけるエクジソン濃度は段階的に上昇してピークに達し、その後、下降する(Hanaoka and Ohnishi, 1974)。本研究の結果はこのエクジソン濃度の時間的変化そのものが、各過程が順序を追って起こることに必要であることを示唆している。
4)高濃度エクジソン注射による鱗毛形成阻害
しかも、エクジソン濃度の段階的変化は、単に諸過程の開始を調節するだけでなく、諸過程の終了も調節しているらしい。本研究において、6.4および12.8μg注射区において鱗毛を分化した個体は各々40%個体、30%個体と少数であり、また鱗毛を分化した個体においても、形成された鱗毛量は少量であった。この事は、高濃度エクジソンは、鱗毛形成を阻害することを示唆している。
同じことは、他の鱗翅目昆虫を用いた腹部表皮の変態の研究でも示されている。休眠中の蛹や分離腹部に脱皮ホルモンを投与すると、成虫分化を誘導することができるが、それが過剰に投与された場合には、鱗毛を持たない成虫に発生する(Kobayashi et.al. 1967, Sakurai and Hasegawa 1969, Judy and Gilbert 1970, Williams 1970, Sedlak and Gilbert 1976)。すなわち、腹部表皮においても、高濃度エクジソンは鱗毛の分化を阻害する。
したがって、鱗毛形成は低濃度エクジソンにより開始・促進されるだけでなく、高濃度エクジソンにより阻害される。このことは、鱗毛細胞の分化や鱗毛突起形成がエクジソン濃度の低い蛹初期に起こり、エクジソン濃度が高い中期には終了する事実とよく一致する。すなわち、変態中のエクジソン濃度の段階的変化は、変態中の各過程の開始と終了の両方を調節することにより、諸過程の順序だった進行を調節していると考えられる。
謝辞
本研究を行うに当たり、有益なご助言をいただき、また一般教育部生物学教室で卒論を作成することを快諾してくださった蚕桑生化学研究室の中島誠教授に感謝します。また、この1年間の研究においていろいろお世話になった一般教育部小原嘉明教授に厚くお礼を申し上げます。
引用文献
Agui,N. and Fukaya,M (1973) Effect of moulting hormones and prothoracic glands on the development of wing disks of cabbage armyworm (Mamestra brassicae) in vitro. Appl. Entmol. Zool., 8(2): 73-82.
Dutkowski,A.B. and Oberlander,H. (1973) The infuluence of larval fat body on wing disc development in vitro. J. Insect Physiol, 19: 2155-62.
Dutkowski,A.B. and Oberlander, H. (1974) Interactions between β-ecdysone and fat body furing wing disc development in vitro. J.Insect Physiol, 20: 743-49.
Hanaoka,K. and Ohnishi,E. (1974) Changes in ecdysone titer during pupal-adult development in the silkworm, Bombyx mori. J. Insect. Physiol. 20: 2375-84.
池田栄太郎 (1913)蚕体解剖生理論.東京:明文堂、東京.
Judy,K.J. and Gilbert,L.I. (1970) Effect of juvenile hormone and moulting hormone on rectal pad development in Hyalophora cecropia. J. Morph. 131: 301-13.
Kobayashi,M. (1967) The molting hormone activity of ecdysterone and inokosterone isolated from Achyranthis radix. J. Insect Physiol. 13: 1395-99.
Sakurai,H. and Hasegawa,K. (1969) Response of isolated pupal abdomens of silkworms, Bombyx mori L., to injected Ponasterone A. Appl. Ent. Zool. 4(2): 69-65.
Sedlak,B.J. and Gilbert,L.I. (1976) Epidermal cell development during the pupal-adult metamorphosis of Hyalophora cecropia. Tissue Cell. 8: 637-48.
Ursprung,H. and Nothiger,E. (1972) The Biology of Imaginal Disks. New York: Springer-Verlag.
Wigglesworth,V.B. (1972) The Principles of Insect Physiology. London: Chapman and Hall.
Williams,C.M. (1970) Hormonal interactions between plants and insect. In Sondheimer,E. and Simeone,J.B. (Ed.) Chemical Ecology. Academic Press, New York. pp. 103-32.
(この論文は1979年2月に東京農工大学蚕糸生物学科に提出された卒業論文を一部修正し、「生徒の理科」見本論文として掲載したものである。)